 |
|
 |
|
|
|
 |
|
綾部 真一 |
|
|
|
| 東京大学大学院理学系研究科修士課程修了 |
| 1973年 |
北里大学薬学部助手
この間1982年〜1983年 セントルイス大学医学部博士研究員
|
| 1989年 |
日本大学農獣医学部(現生物資源科学部)専任講師 |
| 1992年 |
日本大学農獣医学部助教授 |
| 1998年 |
日本大学生物資源科学部教授 |
|
|
|
 |
|
リゾビウム科の細菌とマメ科植物が共生してできる根粒内で,窒素分子が窒素化合物に変えられる.共生窒素固定の巧妙な仕組みを理解することは,工業的な窒素肥料の生産に伴う莫大なエネルギー消費と環境負荷を克服する21世紀型農業への重要なアプローチである.
最近の十数年に,根粒菌のマメ科植物への感染の初期のメカニズムが解明されてきた.マメ科植物から分泌されるフラボノイドが化学シグナルとして根粒菌のnod遺伝子群を活性化し,Nod因子が合成される。Nod因子は根粒菌側の化学シグナルとなって,植物に根粒菌を受け入れ根粒を形成する準備を行わせる.
Nod因子は根粒菌の種によって修飾が異なっている。このような化学シグナルによって,マメ科植物と根粒菌の共生に種レベルでの特異性が保証され,生態学的に効率的なシステムが成立している(図1).
植物にNod因子が受容された後の細胞応答や,根粒数の制御機構が現在世界で研究されている.しかし,感染中期から後期の植物微生物相互作用についてはあまり知見がない.また進入してくる微生物を病害菌ではなく共生のパートナーとして認識する機構と生体防御機構の関係は特に興味深い.植物の生体防御の最前線は化学的バリアである.そしてマメ科植物の代表的な抗菌物質は,共生初期の化学シグナルと共通のフラボノイドに属する.
私たちは根粒菌との共生に働くマメ科植物の因子や共生と防御を制御する機構を,遺伝子レベルから化学物質レベルに至るまで明らかにしようと次のような研究を行っている.
1.マメ科モデル植物ミヤコグサのアクティベーションタギングラインの確立と共生関連遺伝子の単離:ミヤコグサは2倍体のマメ科雑草で,ゲノムサイズが小さく実験室内で簡単に栽培できるので,マメ科のモデル植物として広く利用されており,ゲノム解読も進んでいる.私たちはミヤコグサの効率的な形質転換法を開発し,ゲノム中にエンハンサーのタグ(目印・荷札)を付けた.これによってミヤコグサは機能獲得型の変異を起こし,その原遺伝子の同定が可能になる.これまでに約3000のタギングラインを作製し,バイオリソースとして広く研究者の役に立てる準備も始めている.また窒素栄養飢餓症状を示したり根粒細菌との共生に異常の見られる株が得られ,その変異の原遺伝子の解明を進めている(図2).
2.マメ科に特有なイソフラボノイド生合成経路の解明:イソフラボノイドはマメ科植物の生体防御の主役であり,根粒菌への共生シグナルともなる.これまでにフラボノイドの共通前駆体からイソフラボノイド構造を構築する生合成酵素とその遺伝子を世界に先駆けて同定した.また珍しい分子内転位反応によってイソフラボノイド骨格をつくるシトクロムP450のタンパク質構造と反応性,分子進化に関し詳細な研究を行っている.
3.ミヤコグサのトリテルペノイド生合成酵素の網羅的解析:トリテルペノイドも植物独自の多様性に富む成分であり,植物が環境に適応するためにその機能を利用していると考えられる.鎖状前駆体から環状トリテルペンをつくる環化酵素の遺伝子をミヤコグサから多数クローニングし,機能を解明した.また遺伝子構造をもとに植物が多様な構造を生合成できるようになった過程を推察した.
今後アクティベーションタギングラインから得られた変異株の原遺伝子を明らかにし,またフラボノイドとトリテルペノイドの合成遺伝子やP450による形質転換植物を作製して,化学因子の共生と防御での働きを一層明らかにして行く予定である.
|
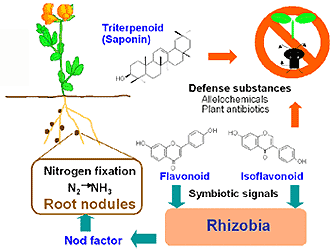 |
| 図1.マメ科植物の化学因子と環境生物の相互作用.フラボノイドやイソフラボノイドが根粒細菌に化学シグナルとして働きかけ,生じたNod因子がマメ科植物に根粒をつくらせるように働きかける.根粒中では窒素固定が行われ,窒素栄養が産み出される.一方,植物のトリテルペノイドやイソフラボノイドは植物がテリトリーを確保したり病原微生物から身を守るのにも働いている. |
|
|
 |
 |
図2.ミヤコグサのアクティベーションタギングラインから単離された変異体候補
A:根粒細菌との共生変異体候補,左から野生型,根粒数の減少,無効根粒着生
B:窒素栄養変異体候補,左から野生型,変異体候補 |
|
|
|
|
|
 |
|
| 1 |
| Shimada N, Aoki T, Sato S, Nakamura Y,
Tabata S, Ayabe S. A cluster of genes encodes
the two types of chalcone isomerase involved
in the biosynthesis of general flavonoids
and legume-specific 5-deoxy(iso)flavonoids
in Lotus japonicus. Plant Physiology 131:
941-951, 2003. |
|
| 2 |
| Ayabe S, Akashi T, Aoki T. Cloning of cDNAs
Encoding P450s in the Flavonoid/Isoflavonoid
Pathway from Elicited Leguminous Cell Cultures,
In Methods in Enzymology, Cytochrome P450,
Part C (Eds, Johnson, E. F., Waterman, M.
R.), pp. 360-369, Academic Press, San Diego,
2002. |
|
| 3 |
| Aoki T, Kamizawa A, Ayabe S. Efficient
Agrobacterium-mediated transformation of
Lotus japonicus with reliable antibiotic
selection. Plant Cell Reports 21: 238-243,
2002. |
|
| 4 |
| Sawada Y, Kinoshita K, Akashi T, Aoki T,
Ayabe S. Key amino acid residues required
for aryl migration catalyzed by the cytochrome
P450 2-hydroxyisoflavanone synthase. Plant
Journal 31: 555-564, 2002. |
|
| 5 |
| Shimada N, Akashi T, Aoki T, Ayabe S. Induction
of isoflavonoid pathway in the model legume
Lotus japonicus: molecular characterization
of enzymes involved in phytoalexin biosynthesis.
Plant Science 160: 37-47, 2000. |
|
|
|
|
 |
|
|
| Copyright 2005 Nihon University
All Rights Reserved. |
|
|
|

