
<メジナとクロメジナ>
メジナ属魚類は、世界で16種が知られている。日本近海にはこのうちメジナGirella punctata、クロメジナG. leoninaおよびオキナメジナG. mezinaの3種が生息しており、その生態は類似している(Yagishita
and Nakabo, 2000; 2003)。特に、メジナおよびクロメジナは非常に似通った外見的特徴をもち、釣りの対象魚として最もポピュラーな魚の一部である。両種は太平洋側では同所的に分布しているものの、日本海側では主にメジナのみが生息するとされ、生態学的に非常に興味深い魚種である。本研究では、分子生物学的手法を利用してメジナおよびクロメジナの生態および進化について調べている。
1.分子生物学的手法を用いるメジナおよびクロメジナの種判別法の確立
両種の外見的特徴からの判別法は報告されているが、稚仔魚および幼魚では計数形質が未分化であるため困難である。そこで本研究では、部分ミトコンドリアDNA(mtDNA)を対象に分子生物学的手法を用いて両種の判別法の確立を目的とした。
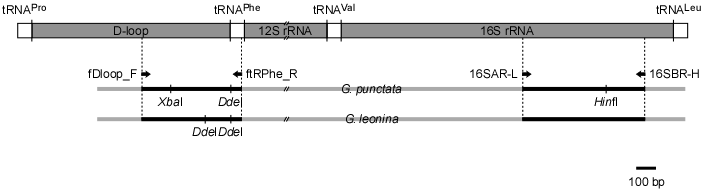
|
図1.メジナおよびクロメジナ部分mtDNAの模式図および種判別に使用可能なプライマーおよび制限酵素認識部位.矢印はPCRに用いるプライマーの位置と向きを示す.G. punctata,メジナ;G. leonina,クロメジナ. |
まず、形態的特徴から確実にメジナおよびクロメジナと判別できる試料を採取し、Yagishita and Nakabo (2000)の報告にもとづき、背鰭棘数と軟条および臀鰭軟条数、側線鱗数などの計数形質からメジナとクロメジナを判別した。その後、筋肉あるいは肝臓から全DNAを抽出し、mtDNAのD-loop領域および16S rRNA遺伝子領域を特異的に増幅するプライマーを用いてPCRを行った(図1)。増幅産物をダイレクトシーケンス法により塩基配列を決定した後、得られた塩基配列から種特異的な制限酵素サイトを検索し(図1)、RFLP解析を行った。
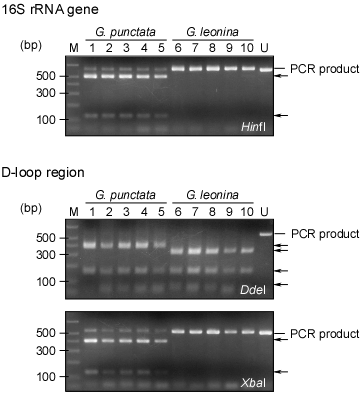 |
|
図2.PCR-RFLP法によるメジナおよびクロメジナの種判別結果.レーンM:分子サイズマーカー;レーン1〜5:メジナ(G. punctata);レーン6〜10:クロメジナ(G. leonina);レーンU:未処理PCR産物.16S rRNA遺伝子にはHinfI、D-loop領域にはDdeIおよびXbaIを用いて判別を行った.矢印は制限酵素処理によって生成されたDNA断片を指す.両種のバンドパターンが明確に異なることが分かる.また、これら制限酵素を用いた場合に予想されるDNA断片は図1に示した模式図と一致する. |
RFLP分析の結果、メジナおよびクロメジナのバンドパターンが明確に異なった(図2)。日本各地から採取した109個体のメジナおよびクロメジナを分析した結果、95個体がメジナ、14個体がクロメジナであることが明らかとなり、この結果は、計数形質をもとに分類した結果と完全に一致した。
本研究の結果は、学術雑誌ICES Journal of Marine Scienceで公表済みである。
2.メジナおよびクロメジナの種分化過程の推定
上述の通り、メジナおよびクロメジナは世界に16種とされるメジナ属魚類の中でも日本近海にのみ生息し、互いにきわめて近縁かつ同所的に分布しているにもかかわらず、それぞれの種を維持している。両種の成り立ちは、生物進化を考える上で非常に興味深い。本研究では、mtDNAにコードされる12Sおよび16S
rRNA遺伝子の全塩基配列を対象に系統解析を行い、メジナおよびクロメジナの分岐年代を推定するとともに、両種の種分化過程を推定した。
メジナ、クロメジナおよびオキナメジナ試料から全DNAを抽出し、完全長の12Sおよび16S
rRNA遺伝子配列を含む部分mtDNAを増幅するPCRを行った。増幅したDNA断片の塩基配列を決定後、Inoue
et al. (2005)のデータをもとにベイズ法を用いて分岐年代を算出した。その結果、図3に示すように、オキナメジナとメジナ/クロメジナの先祖型が分岐したのが730万年前、メジナとクロメジナの先祖型が分岐したのが600万年前と算出された。
| 図3.メジナ属魚類の分岐年代.分岐年代は、Elopomorpha-Clupeocephalaの分岐が2億6500万年前とのデータ(Inoue
et al. 2005)をもとに、12Sおよび16S rRNA遺伝子の全塩基配列を対象としてベイズ法により算出した.系統樹のノード部分の白いバーは、分岐年代の標準偏差を示す. |
|
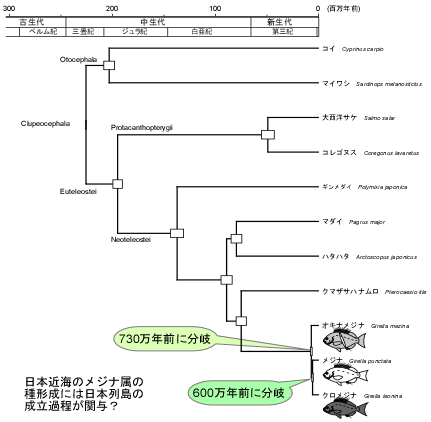 |
メジナ属種間の分岐年代をもとに、図4に示すような仮説を立てた。まず、古日本列島付近に生息していたメジナ属の祖先型が、約700万年前に起きたとされるフィリピン海プレートの沈み込みに伴う琉球弧の隆起により、現在の東シナ海付近と太平洋側に分断された。このとき現在の東シナ海付近の海域に隔離されたメジナ属の祖先型がオキナメジナの祖先型へ、また太平洋側に残ったグループがメジナおよびクロメジナの共通の祖先型へと分化していったと考えられる。さらに、太平洋側に残されたメジナおよびクロメジナの共通の祖先型は、その後、約600万年前の古日本列島付近の造山運動に伴い現在の日本海と太平洋側に分断され、日本海に隔離されたグループがメジナへ、太平洋側に取り残されたグループがクロメジナへと分化したと推測される。その後、日本海のメジナが太平洋側に進出することで、日本列島の太平洋側ではメジナとクロメジナが共存しているのに対し、水温等の環境要因によりクロメジナは日本海側へ分布域を広げることができず、現在の両種の分布域が異なっているものと考えられる。
本研究の結果は、学術雑誌Organisms, Diversity & Evolutionで公表済みである。
3.引用文献
Taira, A., 2001. Tectonic evolution of the Japanese Island arc system. Annu. Rev. Earth Planet. Sci. 29, 109-134.
Yagishita, N., Nakabo, T., 2000. Revision of the genus Girella (Girellidae) from East Asia. Ichthyol. Res. 47, 119-135.
Yagishita, N., Nakabo, T., 2003. Evolutionary trend in feeding habits of Girella (Perciformes: Girellidae). Ichthyol. Res. 50, 358-366.
Inoue, J.G., Miya, M., Venkatesh, B., Nishida, M., 2005. The mitochondrial genome of Indonesian coelacanth Latimeria menadoensis (Sarcopterygii: Coelacanthiformes) and divergence time estimation between the two coelacanths. Gene 349, 227-235.
Home
|
